重慶醫科大(dà)學附屬第一醫院鄧亮老師的(de)課題組在 CLINICAL AND TRANSLATION MEDICINE 雜(zá)志上發表了(le)研究論文 “Runx2 activates hepatic stellate cells to promote liver fibrosis via transcriptionally regulating Itgav expression”, 研究了(le) Runx2 在肝星狀細胞激活和(hé)肝纖維化(huà)中的(de)作用(yòng)。肝星狀細胞(HSC)被認爲是肝纖維化(huà)的(de)潛在治療靶點。轉錄因子 Runx2 與非酒精性脂肪性肝病的(de)發展有關,在 HSC 激活和(hé)肝纖維化(huà)中的(de)具體作用(yòng)暫不清楚。在這(zhè)項研究中,研究人(rén)員(yuán)發現 Runx2 在不同病因的(de)人(rén)類肝纖維化(huà)中顯著上調,在小鼠的(de)纖維化(huà)過程中也(yě)發現了(le)類似的(de)現象,并且其主要在活化(huà)的(de) HSC 中表達。研究人(rén)員(yuán)通(tōng)過構建一種在 HSC 細胞中特異敲除 Runx2 基因的(de)小鼠,使用(yòng)該小鼠進行肝纖維化(huà)誘導,可(kě)發現 HSC 特異敲除 Runx2 基因的(de)小鼠可(kě)以顯著的(de)減輕被誘導小鼠的(de)肝纖維化(huà)過程。進一步的(de)研究發現 Runx2 可(kě)以促進 HSC 的(de)活化(huà)和(hé)增殖,在 HSC 中敲除 Runx2 可(kě)以抑制這(zhè)些作用(yòng)。研究人(rén)員(yuán)又通(tōng)過 RNA-seq and ChIP-seq 技術,分(fēn)析發現 Runx2 可(kě)以促進 Itgav 基因的(de)表達而抑制 Itgav 則可(kě)以減弱 Runx2 誘導的(de) HSC 活化(huà)和(hé)肝纖維化(huà)。因此,此次研究确認了(le) Runx2 在肝纖維化(huà)過程中活化(huà) HSC 的(de)重要作用(yòng),它可(kě)能是肝纖維化(huà)研究和(hé)治療的(de)一個(gè)非常有潛力的(de)靶點。
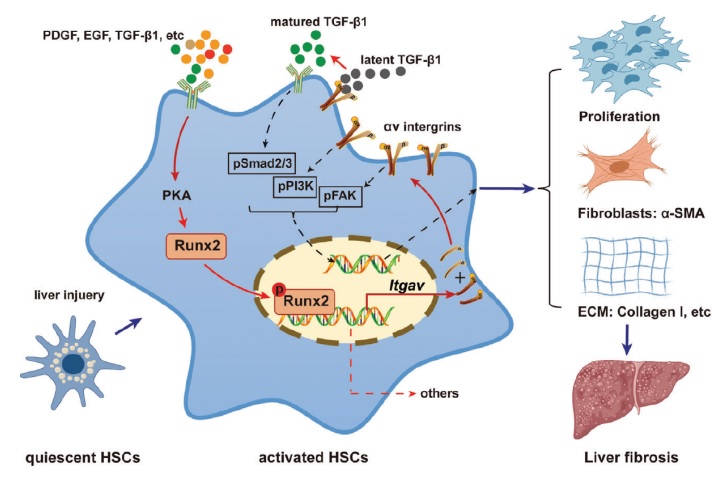
背景和(hé)目标
肝星狀細胞(HSC)作爲肝纖維化(huà)的(de)關鍵環節,被認爲是肝纖維化(huà)的(de)潛在治療靶點。已有研究表明(míng),轉錄因子 Runx2 與非酒精性脂肪肝的(de)發展有關,而其在 HSC 激活和(hé)肝纖維化(huà)中的(de)具體作用(yòng)仍然沒有明(míng)确。
方法和(hé)結果
在這(zhè)項研究中,研究人(rén)員(yuán)發現 Runx2 的(de)表達在不同病因所形成的(de)人(rén)類肝纖維化(huà)中都顯著上調。在纖維化(huà)過程中 Runx2 在小鼠肝髒中的(de)表達也(yě)逐漸升高(gāo),并且 Runx2 主要在活化(huà)的(de) HSC 中表達。在 HSC 中 Runx2 的(de)敲除顯著減輕了(le) CCl4, DDC or MCD 誘導的(de)肝纖維化(huà),而通(tōng)過對(duì)小鼠的(de)肝進行 HBAAV-Runx2 或 VA-Lip-Runx2 的(de)注射過表達 Runx2 加劇了(le) CCl4 誘導的(de)肝纖維變性。體外的(de)分(fēn)析表明(míng) Runx2 促進了(le) HSC 的(de)活化(huà)和(hé)增殖,而在 HSC 中敲低 Runx2 則抑制了(le)這(zhè)些作用(yòng)。通(tōng)過 RNA-seq 和(hé) Runx2-ChIP-seq 的(de)分(fēn)析表明(míng) Runx2 可(kě)以通(tōng)過結合啓動子來(lái)促進 Itgav 基因的(de)表達。阻斷 Itgav 則可(kě)減弱 Runx2 誘導的(de) HSC 活化(huà)和(hé)肝纖維化(huà)。此外,研究還(hái)發現細胞因子 TGF-β1, PDGF and EGF 可(kě)以通(tōng)過蛋白激酶 A(PKA)來(lái)促進 HSC 中 Runx2 的(de)表達和(hé)核轉位。
結論
轉錄因子 Runx2 在肝纖維化(huà)過程中通(tōng)過轉錄調節 Itgav 的(de)表達,對(duì) HSC 的(de)活化(huà)至關重要,它可(kě)能是肝纖維化(huà)的(de)一個(gè)比較有前途的(de)治療靶點。
1. 在肝纖維化(huà)進展的(de)過程中 Runx2 的(de)表達逐漸增加爲了(le)研究 Runx2 在肝纖維化(huà)中的(de)作用(yòng),研究人(rén)員(yuán)首先在 GEO 數據庫中探索了(le) Runx2 的(de)表達。在由不同病因引起的(de)肝硬化(huà)患者中 Runx2 的(de)表達要高(gāo)得(de)多(duō),如酒精性肝病、病毒性肝炎和(hé) NAFLD(圖 1 A)。研究人(rén)員(yuán)還(hái)發現 Runx2 的(de)表達水(shuǐ)平在晚期纖維化(huà)患者中較高(gāo),而在早期纖維化(huà)患者中較低,這(zhè)表明(míng) Runx2 的(de)表達在肝纖維化(huà)的(de)進展過程中會增加。接著(zhe),研究人(rén)員(yuán)從非纖維化(huà)和(hé)肝硬化(huà)患者的(de)身上收集人(rén)類肝組織,結果也(yě)顯示 Runx2 的(de)蛋白和(hé) mRNA 水(shuǐ)平在肝硬化(huà)患者的(de)肝組織中顯著上調(圖 1 B and C)。組織學分(fēn)析馬松染色顯示肝硬化(huà)組織中膠原嚴重沉積,同時(shí) Runx2 and α-SMA(活化(huà) HSC 的(de)标志物(wù))表達顯著增加。在纖維索處 Runx2 也(yě)呈陽性染色(圖 1 D)。在 GEO 數據庫和(hé) CCl4 引起的(de)小鼠纖維化(huà)模型中,在纖維化(huà)的(de)肝組織中也(yě)觀察到了(le)類似的(de) Runx2 表達上調的(de)結果(圖 1 E-H)。更重要的(de)是,研究人(rén)員(yuán)發現 Runx2 and α-SMA 在高(gāo)脂飲食誘導的(de)非酒精性脂肪性肝炎(NASH)相關的(de)肝纖維化(huà)中均顯著增加(圖 1 H)。爲了(le)進一步研究 Runx2 在肝纖維化(huà)過程中的(de)表達趨勢,研究人(rén)員(yuán)用(yòng) CCl4 處理(lǐ)小鼠 1 周和(hé) 6 周以誘導肝的(de)損傷和(hé)纖維化(huà),并監測 Runx2 的(de)表達。研究發現,随著(zhe)肝損傷的(de)持續,膠原沉積逐漸增加,在受損肝髒中 α-SMA and Runx2 的(de)表達輕度上調,但在纖維化(huà)肝髒中明(míng)顯增加,這(zhè)與 GEO 數據庫的(de)分(fēn)析一緻。有趣的(de)是,組織學染色結果表明(míng) Runx2 的(de)陽性染色主要分(fēn)布在肝組織的(de)纖維索和(hé)間隔中,與 α-SMA 陽性細胞的(de)分(fēn)布一緻(圖 1 D and H)。總之,這(zhè)些發現表明(míng) Runx2 的(de)表達在肝纖維化(huà)過程中逐漸增加,并在肝組織中表現出潛在的(de)細胞特異性表達的(de)特征。
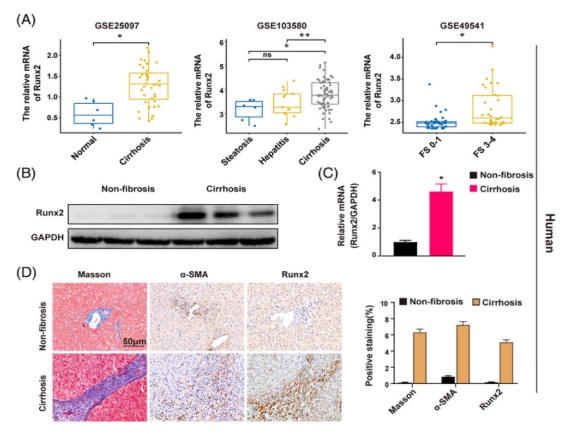
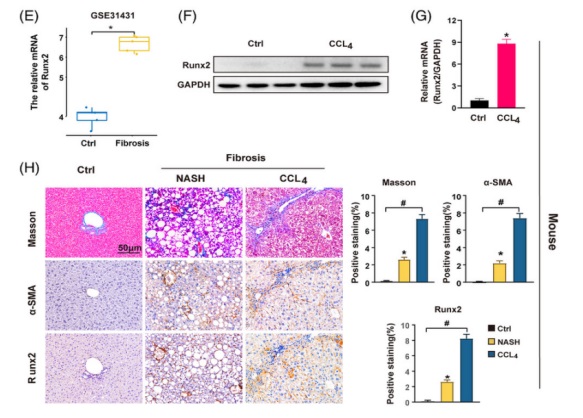
fig.1
由于 Runx2 主要在肝間質細胞中表達,與 α-SMA 的(de)表達一緻,研究人(rén)員(yuán)在纖維化(huà)的(de)小鼠肝髒中用(yòng) α-SMA(活化(huà)的(de) HSC 标記物(wù)), CD31(内皮細胞标記物(wù)) 和(hé) F4/80(庫普弗細胞标志物(wù))對(duì) Runx2 進行共染,以評估 Runx2 在肝髒中表達空間的(de)特異性。正如預測的(de)那樣,僅在 Runx2 和(hé) α-SMA 之間觀察到共表達的(de)現象,這(zhè)在人(rén)類肝硬化(huà)的(de)肝組織中得(de)到了(le)進一步證實(圖 2 A)。此外,通(tōng)過比較靜态的(de)和(hé)從 CCl4 處理(lǐ) 4 周的(de)小鼠中分(fēn)離的(de)活化(huà)的(de) HSC 之間的(de) Runx2 表達,發現活化(huà) HSC 中 Runx2 的(de)表達顯著增加(圖 2 B)。值得(de)注意的(de)是,單細胞 RNA 測序分(fēn)析顯示,與早期培養的(de)原代 HSC 相比 Runx2 在肌成纖維細胞(分(fēn)化(huà)良好的(de) HSC)中高(gāo)度表達和(hé)分(fēn)布,這(zhè)與肌纖維細胞标記物(wù)(α-SMA and Col1a1)一緻,這(zhè)進一步支持 Runx2 主要存在于活化(huà)的(de) HSC 中(圖 2 C)。最重要的(de)是,在 HSC 的(de)激活過程中 Runx2 的(de) mRNA 和(hé)蛋白表達以時(shí)間依賴性的(de)方式進行上調,這(zhè)與纖維化(huà)基因的(de)表達有關,包括 α-SMA, collagen I and TGF-β1(圖 2 D and E)。同時(shí),研究人(rén)員(yuán)還(hái)觀察到 Runx2 在體外表現出過度的(de)核轉位并伴有 HSC 活化(huà),這(zhè)意味著(zhe) Runx2 同時(shí)開始了(le)其轉錄功能(圖 2 F)。總之,這(zhè)些結果表明(míng) Runx2 在活化(huà)的(de) HSC 中特異性表達,并可(kě)能在調節 HSC 活化(huà)中發揮作用(yòng)。
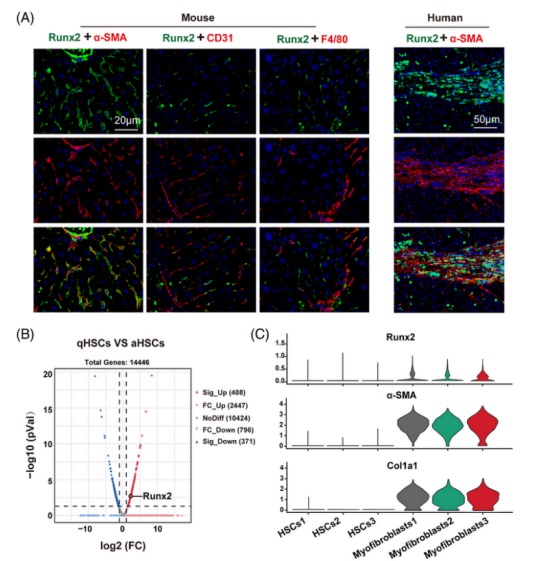
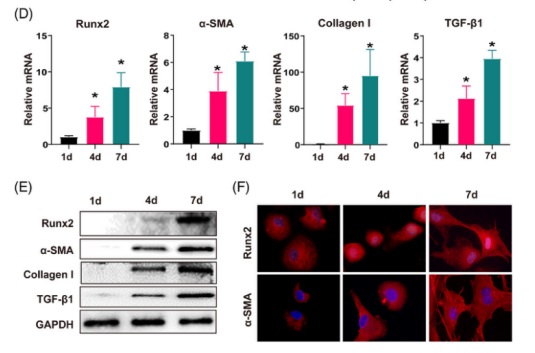
fig.2
3. HSC 特異性敲低 Runx2 可(kě)以減輕 CCl4, DDC and MCD 誘導的(de)小鼠肝纖維化(huà)爲了(le)确定 Runx2 在體内肝纖維化(huà)過程中的(de)作用(yòng),研究人(rén)員(yuán)首先使用(yòng)慢(màn)病毒介導的(de) shRNA 在 CCl4 和(hé)橄榄油對(duì)照(zhào)誘導的(de)小鼠中敲低了(le) Runx2 數據表明(míng)敲低 Runx2 減輕了(le) CCl4 誘導的(de)肝纖維化(huà)。接下(xià)來(lái),研究人(rén)員(yuán)又通(tōng)過使用(yòng) PDGFRβ-Cre(HSC 特異性基因) 小鼠來(lái)構建出了(le) HSC 特異性敲除 Runx2 基因的(de)小鼠。然而,純合子(Runx2ff;PDGFRα-Cre)在出生後立即死亡,因此 Runx2 半敲除的(de)雜(zá)合子小鼠(Runx2f+;PDGFRβ-Cre or Runx2△+HSC)被用(yòng)于進一步的(de)體内研究(圖 3 A)。結果表明(míng),在 CCl4 誘導的(de)纖維化(huà)組中 Runx2 的(de)敲低降低了(le)促纖維化(huà)基因,包括 TGF-β1, collagen I and α-SMA 的(de) mRNA 和(hé)蛋白水(shuǐ)平的(de)表達。盡管它們的(de) mRNA 表達水(shuǐ)平在用(yòng)橄榄油處理(lǐ)的(de) Runx2△+HSC 小鼠中輕度下(xià)降,但這(zhè)些基因的(de)蛋白質水(shuǐ)平沒有顯示出顯著的(de)變化(huà)(圖 3 B and C)。此外,敲低 Runx2 在組織學上顯著減輕了(le) CCl4 誘導的(de)肝纖維化(huà),減少了(le) I 型膠原的(de)沉積和(hé) HSC 的(de)活化(huà)(圖 3 D)。此外,研究人(rén)員(yuán)還(hái)建立了(le)DDC or MCD 誘導的(de)肝纖維化(huà)小鼠模型,以評估 Runx2 在其他(tā)病因引起的(de)肝纖維化(huà)中的(de)作用(yòng)。通(tōng)過 H&E 染色證實 DDC 或 MCD 成功在小鼠中誘導了(le)膽汁淤積症和(hé) NASH, 另外,與對(duì)照(zhào)組相比 Runx2 敲除小鼠的(de)纖維化(huà)和(hé) HSC 的(de)激活水(shuǐ)平都顯著降低(圖 3 E)。類似地 Runx2 的(de)敲降也(yě)降低了(le) DDC and MCD 誘導的(de)纖維化(huà)肝組織中 I 型膠原和(hé) α-SMA 的(de) mRNA 和(hé)蛋白質的(de)表達。此外,由于肝細胞(HC)損傷是肝纖維化(huà)的(de)一個(gè)組成部分(fēn),研究人(rén)員(yuán)通(tōng)過使 Runx2ff 小鼠與 Alb-Cre 小鼠來(lái)進行繁育也(yě)構建了(le) HC 特異性敲除 Runx2 的(de)小鼠(Runx2△/△HCs)來(lái)驗證在 HC 中的(de) Runx2 敲除是否會影(yǐng)響肝纖維化(huà)的(de)進展。不出所料 Runx2 的(de) HC 特異性敲除并不會影(yǐng)響 CCl4 誘導的(de)肝纖維化(huà)。總之,這(zhè)些發現表明(míng) HSC 中 Runx2 的(de)敲除減輕了(le) CCl4, DDC and MCD 誘導的(de)肝纖維化(huà)。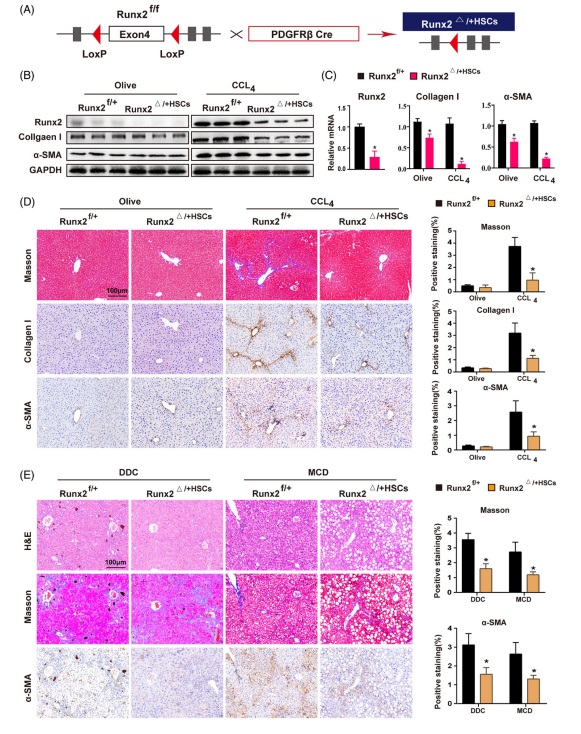
fig.3
4. Runx2 的(de)過表達加劇了(le) CCl4 誘導的(de)肝纖維化(huà)爲了(le)進一步證實 Runx2 在肝纖維化(huà)中的(de)作用(yòng),研究人(rén)員(yuán)通(tōng)過注射 HBAAV-Runx2 三周來(lái)構建 Runx2 過表達小鼠,然後又用(yòng)橄榄油和(hé) CCl4 來(lái)處理(lǐ)小鼠 4 周(圖 4 A)。随後,分(fēn)離原代 HSC 和(hé) HC 并檢測 Runx2 基因 mRNA 的(de)表達,以弄清過表達質粒在肝不同細胞群間的(de)遞送和(hé)表達效率。結果顯示,與 HSC 相比,大(dà)多(duō)數 HBAAV-Runx2 被轉染到了(le) HC 中。然而,由于 Runx2如前所述并沒有影(yǐng)響 HC, 因此在某種程度上研究人(rén)員(yuán)成功地構建了(le)在 HSC 中過表達 Runx2 的(de)小鼠。在使用(yòng) CCl4 處理(lǐ)後,結果發現,與對(duì)照(zhào)組相比,在 HBAAV-Runx2 過表達小鼠中 Runx2, collagen I and α-SMA 的(de) mRNA 和(hé)蛋白的(de)表達都顯著增加(圖 4 B and C)。在組織學上,與對(duì)照(zhào)組相比,過表達 HBAAV-Runx2 組小鼠的(de)肝髒結構表現出廣泛的(de)結構紊亂和(hé)相鄰血管間顯著的(de)間隙(圖 4 D)。在 HBAAV-Runx2 小鼠中 I 型膠原和(hé) α-SMA 細胞的(de)陽性染色也(yě)增加,表明(míng)纖維化(huà)嚴重,大(dà)量 HSC 被激活(圖 4 D)。此外,研究人(rén)員(yuán)發現在橄榄油處理(lǐ)的(de)小鼠中 Runx2 的(de)過表達觸發了(le) GFAP 陽性細胞(靜态 HSC)的(de)增殖,但不影(yǐng)響膠原沉積, HSC 的(de)活化(huà), I 型膠原和(hé) α-SMA 的(de) mRNA 和(hé)蛋白的(de)表達(圖 4 D),這(zhè)表明(míng) Runx2 在 HSC 中的(de)過表達不能觸發自發纖維化(huà)或 HSC 活化(huà)。此外,研究發現,通(tōng)過注射攜帶 Runx2(VA-Lip-Runx2) 或 VA-Lip-Ctrl 質粒的(de)脂質體,在 HSC 中特異性過表達 Runx2 增強了(le) CCl4 誘導的(de)小鼠肝纖維化(huà)。綜上所述,這(zhè)些結果表明(míng) Runx2 的(de)過表達會加劇 CCl4 誘導的(de)肝纖維化(huà)。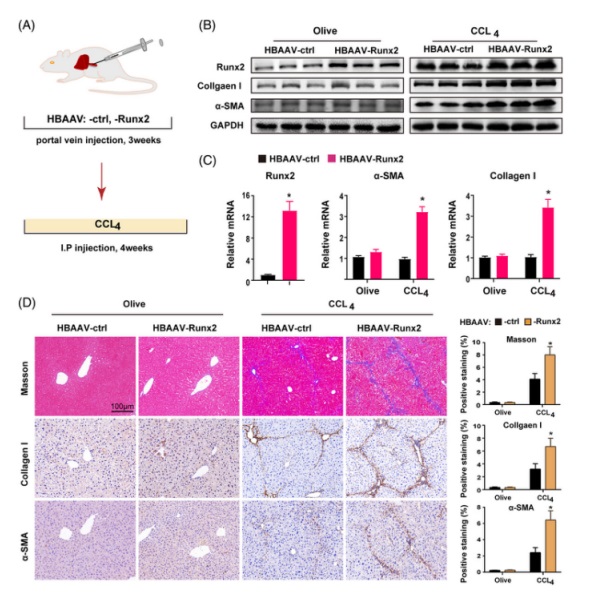
fig.4
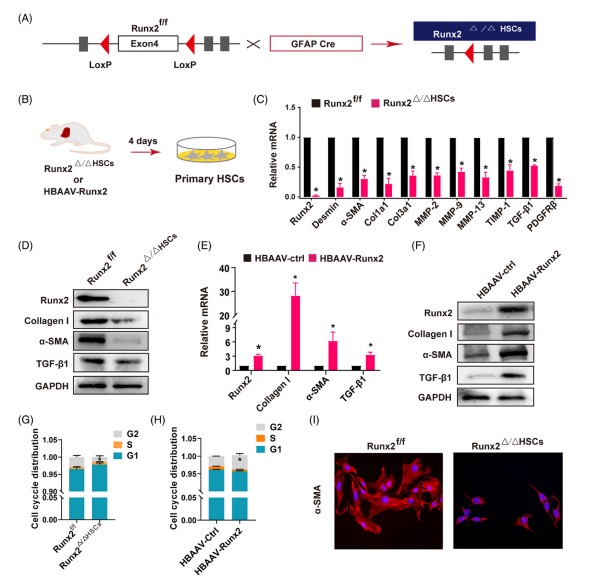
5. Runx2 在體外調節 HSC 的(de)激活由于 Runx2f/f 小鼠與 PDGFRβ-Cre 小鼠隻能培育出 Runx2 半敲除的(de)雜(zá)合子小鼠(Runx2△/+HSC);研究人(rén)員(yuán)利用(yòng) GFAP-Cre(肝中 HSC 的(de)另一種标記物(wù))小鼠與 Runx2f/f 小鼠雜(zá)交,成功獲得(de) Runx2 敲除純合子小鼠(Runx2△/△HSC)(圖 5 A)。考慮到 GFAP-Cre 也(yě)可(kě)以标記膽管細胞,所以 Runx2△/△HSC 小鼠在本次研究中主要被用(yòng)于進行體外實驗。爲了(le)進一步評估 Runx2 在 HSC 激活中的(de)作用(yòng),從 Runx2△/△HSC 和(hé) HBAAV-Runx2 小鼠中分(fēn)離出原發性 HSC 并培養 4 天(圖 5 B)。結果發現在 Runx2△/△HSC 小鼠缺失 Runx2 基因的(de) HSC 中 HSC 标記基因和(hé)纖維化(huà)基因,如 desmin, α-SMA, Col1a1, Col3a1, MMP2, MMP9, MMP13, TIMP-1, TGF-β1 和(hé) PDGFRβ 的(de) mRNA 表達都降低了(le)(圖 5 C),并且 α-SMA, collagen I and TGF-β1 的(de)蛋白表達也(yě)降低了(le)(圖 5 D),然而在 Runx2 過表達小鼠的(de) HSC 中,纖維化(huà)基因如 α-SMA, collagen I and TGF-β1 的(de) mRNA 和(hé)蛋白的(de)表達都提高(gāo)了(le)(圖 5 E and F)。此外 Runx2 缺失抑制了(le) α-SMA 的(de)表達,并在體外維持了(le) HSC 靜态的(de)表型(圖 5 I)。接著(zhe),研究人(rén)員(yuán)進一步使用(yòng)小幹擾 RNA 來(lái)敲低原代 HSC, LX2(人(rén) HSC 細胞系)和(hé) mHSC (小鼠 HSC 細胞系)中的(de) Runx2. 結果顯示 Runx2 缺失降低了(le)所有細胞中 α-SMA mRNA 的(de)表達,而 Runx2 過表達上調了(le) mHSC 細胞中 α-SMA mRNA 的(de)表達。此外,流式細胞分(fēn)析的(de)結果顯示 Runx2 的(de)敲除顯著減少了(le) HSC 的(de) G2 期,表明(míng) HSC 增殖受到抑制,但 Runx2 的(de)過表達會持續導緻 HSC 細胞 S 期的(de)減少和(hé) G2 期的(de)增加,并顯著促進了(le) HSC 在體外的(de)生長(cháng)(圖 5 G),這(zhè)表明(míng) Runx2 是 HSC 增殖的(de)正向調節因子。總之,這(zhè)些發現表明(míng) Runx2 是 HSC 激活和(hé)增殖的(de)重要轉錄因子。
fig.5
6 .Itgav 是 Runx2 在肝纖維化(huà)中的(de)直接下(xià)遊靶點爲了(le)探索 Runx2 調節 HSC 激活的(de)潛在機制,研究人(rén)員(yuán)通(tōng)過使用(yòng)轉染了(le)幹擾 Runx2 表達的(de) siRNA 的(de)原代 HSC 來(lái)進行 RNA 測序(RNA-seq)。根據 KEGG 富集分(fēn)析的(de)結果顯示 Runx2 敲低主要與激活 HSC 的(de)信号通(tōng)路受損有關,包括凋亡、細胞周期、自噬、MAPK and TGF-β 信号通(tōng)路(圖 6 A)。由于 Runx2 在各種生物(wù)過程中起轉錄因子的(de)作用(yòng),所以研究人(rén)員(yuán)對(duì)來(lái)自 HBAAV-Runx2 小鼠的(de) HSC 又進行了(le)染色質免疫沉澱測序(ChIP-seq)分(fēn)析,以确定 Runx2 在活化(huà)的(de) HSC 中的(de)直接靶點。結果顯示 Runx2 與許多(duō) DNA 序列有結合,其中許多(duō)峰位于靶基因的(de)轉錄起始位點附近。根據 KEGG 分(fēn)析發現 Runx2 結合的(de)區(qū)域主要靠近 PI3K-Akt 信号通(tōng)路、代謝通(tōng)路、TGF-β 信号通(tōng)路等相關基因(圖 6 B)。進一步 RNA-seq and ChIP-seq 顯示 Runx2 對(duì) TGF-β 信号通(tōng)路有顯著影(yǐng)響。然而,包括 TGF-β1 在内的(de) TGF-β 配體并不是 Runx2 的(de)直接下(xià)遊因子,這(zhè)表明(míng) Runx2 可(kě)能通(tōng)過不直接介導 TGF-β 配體的(de)表達的(de)方式來(lái)調節 TGF-β 信号通(tōng)路。由于 TGF-β 信号通(tōng)路是調節 HSC 活化(huà)和(hé)肝纖維化(huà)過程中最重要的(de)通(tōng)路之一,整合 TGF-β 通(tōng)路相關基因轉錄組和(hé) ChIP-seq 數據集,獲得(de)了(le) 7 個(gè)常見基因,包括 Itgav, Thbs1, Lox, Src, Spry2, Smad7 和(hé) Npnt(圖 6 C)。其中 Itgav 基因編碼了(le) αv 整合素的(de)主要成分(fēn),而 αv 整合素是調節 TGF-β1 激活和(hé)肝纖維化(huà)進展的(de)關鍵整合素,這(zhè)表明(míng) Runx2 可(kě)能是通(tōng)過 Itgav 信号通(tōng)路來(lái)激活 HSC 的(de)。使用(yòng) ChIP-seq 分(fēn)析發現 Runx2 與 Itgav 轉錄起始位點上遊的(de) Chr2:8372357-83723806 結合,這(zhè)種結合通(tōng)過熒光(guāng)素酶報告基因測定實驗得(de)到了(le)證實(圖 6 D and E)。此外,對(duì)來(lái)自 Runx2△/△HSC, Runx2△/+HSC 和(hé) HBAAV-Runx2 小鼠的(de)肝組織和(hé)原代 HSC 中 Itgav 基因的(de) mRNA 和(hé)蛋白表達進行了(le)檢查,結果表明(míng) Runx2 缺乏或過表達在體外和(hé)體内降低或增加了(le) Itgav 的(de)表達(圖 6 F-I)。鑒于 Itgav 與 ECM 配體結合并激活下(xià)遊激酶,包括 FAK and PI3K. 以及與 TGF-β1 相互作用(yòng)以激活 TGF-β1 信号通(tōng)路。因此,研究人(rén)員(yuán)檢測了(le)來(lái)自 Runx2 敲除和(hé)過表達小鼠原代 HSC 中的(de)磷酸化(huà) FAK, PI3K and Smad2/3 的(de)表達。結果表明(míng),磷酸化(huà) FAK, PI3K and Smad2/3 的(de)表達在 Runx2 過表達的(de) HSC 中增強,而在 Runx2 敲除的(de) HSC 則降低(圖 6 I)。總之,我們的(de)研究結果強烈表明(míng) Runx2 通(tōng)過與 Itgav 啓動子結合來(lái)直接上調 Itgav 的(de)表達,并激活潛在的(de)信号轉導,這(zhè)反過來(lái)又有助于 HSC 的(de)激活和(hé)肝纖維化(huà)的(de)進展。同時(shí),相關分(fēn)析顯示 Runx2 與 Itgav (r=.98), TGF-β1(r=.91) 和(hé) collagen I(r=.97)高(gāo)度相關。且 Itgav 也(yě)與 TGF-β2(r=.87) and collagen I(r=0.94)高(gāo)度相關。這(zhè)些結果表明(míng),這(zhè)些基因之間可(kě)能存在一些功能關系,它們可(kě)能共同調節或參與相似的(de)生物(wù)通(tōng)路。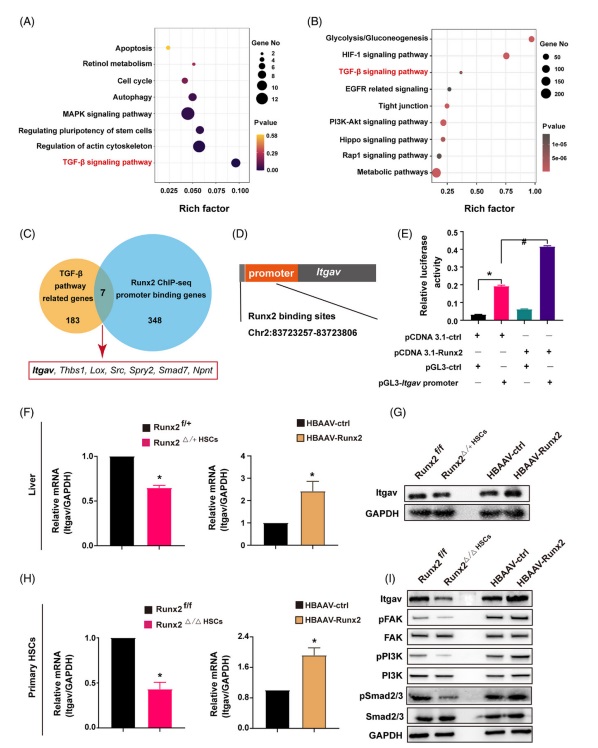
fig.6
7 .抑制 αv 整合素阻斷了(le) Runx2 過表達引起的(de) CCl4 誘導的(de)肝纖維化(huà)加重爲了(le)探索 Itgav 在 Runx2 相關 HSC 激活和(hé)肝纖維化(huà)中的(de)作用(yòng),研究人(rén)員(yuán)首先通(tōng)過在從HBAAV-ctrl 和(hé) HBAAV-Runx2 小鼠中分(fēn)離出的(de)原代 HSC 中使用(yòng)小幹擾 RNA 來(lái)沉默 Itgav 基因,結構顯示 Itgav 敲低顯著阻斷了(le) Runx2 過表達誘導的(de) α-SMA 上調。由于 Itgav 編碼的(de)整合素 α-V 鏈是五種 αV 整合素的(de)主要成分(fēn),其表達變化(huà)将影(yǐng)響所有五種 αV 整合素的(de)功能,因此,研究人(rén)員(yuán)進一步利用(yòng) αv 整合素的(de)小分(fēn)子抑制劑(CWHM-12)來(lái)确定 Itgav 介導的(de) αv 整合素功能是否确實在 HSC 激活和(hé)肝纖維化(huà)進展中處于 Runx2 調節的(de)下(xià)遊(圖 7 A)。結果發現 αv 整合素抑制顯著阻斷了(le) Runx2 過表達引起的(de) α-SMA 上調,這(zhè)與 siItgav 相似(圖 7 B and C)。然後,給 HBAAV-Runx2 或 HBAAV-ctrl 小鼠注射 CCl4 誘導 2 周以構建肝纖維化(huà),然後再給藥 CWHM-12 或 DMSO 處理(lǐ) 2 周(圖 7 D)。結果顯示 CWHM-12 顯著降低了(le) α-SMA 的(de) mRNA 和(hé)蛋白質表達,并阻斷了(le) Runx2 誘導的(de) α-SMA 表達上調,這(zhè)與體外數據一緻(圖 7 E and F)。在組織學檢測中,通(tōng)過膠原沉積(Masson and collagen I 染色)和(hé) α-SMA 染色檢測顯示 CWHM-12 顯著減少了(le)肝纖維化(huà),重要的(de)是抑制 αv 整合素顯著阻斷了(le) Runx2 加重的(de)肝纖維化(huà)(圖 7 G)。研究人(rén)員(yuán)還(hái)通(tōng)過在小鼠中同步靜脈注射攜帶 Itgav siRNA 的(de)脂質體(VA-Lip-siItgav) 和(hé) VA-Lip-Runx2 來(lái)特異性阻斷 Itgav 在 HSC 中的(de)表達,以避免 CWHM-12 可(kě)能潛在的(de)副作用(yòng),結果顯示 siItgav 也(yě)顯著抑制了(le) Runx2 促進的(de)肝纖維化(huà)。因此,研究結果表明(míng) Runx2 主要通(tōng)過 Itgav 激活 HSC 并促進肝纖維化(huà)。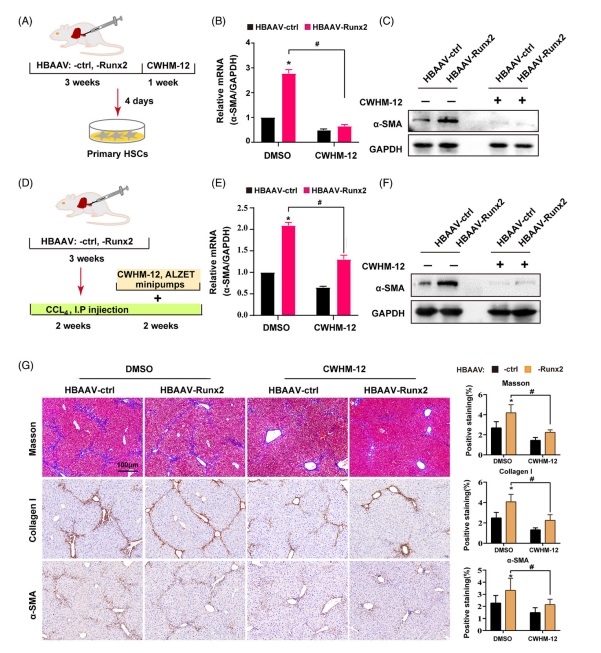
fig.7
8 .在 HSC 中 Runx2 的(de)激活和(hé)核轉位受 PKA 的(de)調節Runx2 從細胞質到細胞核的(de)易位是其轉錄活性所必需的(de)。因此,研究人(rén)員(yuán)也(yě)試圖找出提高(gāo) Runx2 表達并控制其易位進入細胞核的(de)上遊調節信号。研究表明(míng),蛋白激酶 A(PKA)激活 Runx2 并在腸上皮細胞的(de)上皮-間質轉化(huà)和(hé)成骨細胞分(fēn)化(huà)過程中介導其核轉位。此外 PKA 也(yě)是 PDGF, EGF 和(hé) TGF-β1 的(de)常見下(xià)遊激酶,它們是調節 HSC 激活的(de)必需細胞因子。因此,研究人(rén)員(yuán)利用(yòng) TGF-β1, PDGF-BB 或 EGF 來(lái)激活使用(yòng)或不用(yòng) PKA 抑制劑(PKI-6-22)處理(lǐ)的(de)原代 HSC 細胞。結果顯示 TGF-β1, PDGF-BB 和(hé) EGF 都顯著增加了(le) Runx2 的(de)表達,而 PKA 抑制劑則消除了(le)這(zhè)效應(圖 8 A)。此外 Runx2 可(kě)以激活 Itgav 的(de)轉錄,因此也(yě)檢測了(le) PKA 對(duì) Itgav 的(de)影(yǐng)響。結果顯示 TGF-β1, PDGF-BB 和(hé) EGF 顯著增加了(le) Itgav 的(de)表達,但所有這(zhè)些也(yě)都被 PKA 抑制劑消除了(le)(圖 8 A)。此外,研究還(hái)發現 PKA 激活劑(8-Bromo-cAMP)可(kě)以刺激 Runx2 在 HSC 中的(de)核分(fēn)布。相反 PKA 抑制劑在有或沒有 TGF-β1 培養的(de) HSC 中減弱了(le) Runx2 的(de)核轉位(圖 8 B)。免疫熒光(guāng)染色還(hái)表明(míng),用(yòng) PKA 抑制劑處理(lǐ)的(de) HSC 導緻了(le)細胞核中 Runx2 的(de)表達受到了(le)抑制,使得(de) HSC 表型保持更加靜止,而 PKA 激活劑誘導了(le) Runx2 在細胞核中的(de)分(fēn)布更高(gāo)并且使得(de) HSC 活化(huà)(圖 8 C)。總之,這(zhè)些發現表明(míng) TGF-β1, PDGF-BB 或 EGF 通(tōng)過 PKA 激活 Runx2 并促進其核轉位。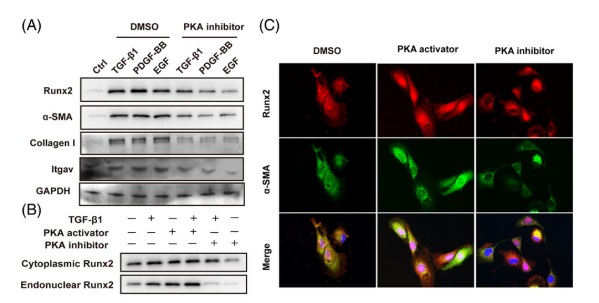
fig.8
本研究中所使用(yòng)的(de) Runx2 CKO 條敲小鼠由北(běi)京唯尚立德提供,該小鼠是在 C57BL/6 的(de)背景下(xià)通(tōng)過 CRISPR/Cas 9 技術構建而成的(de)。北京天糧生物科技有限公司是擁有多(duō)年的(de)基因編輯經驗,已爲數千名科研工作者及工業客戶成功定制基因編輯模型,同時(shí)搭建了(le)完善的(de)基因工程動物(wù)創制平台、動物(wù)表型分(fēn)析和(hé)實驗技術服務平台、動物(wù)模型種源交流平台、實驗動物(wù)繁育平台,歡迎您緻電咨詢。
原文鏈接:https://onlinelibrary.wiley.com/doi/full/10.1002/ctm2.1316
返回列表


 京公網安備 11011402010692号
京公網安備 11011402010692号




